Projects
Cognitive Neuroscience of Aging
The underlying theme to our research focuses on the cognitive neuroscience of aging, wherein we examine the relationship between cognitive processes and cerebral or brain changes in aging (for a review see Dennis & Cabeza 2008). The discipline uses functional neuroimaging techniques such as functional magnetic resonance imagining (MRI) to evaluate age-related functional change within cerebral grey matter during cognitive tasks. Our lab also examines the link between structural and function in aging in order to elucidate the basis for age-related decline in cognitive processes. While our examination includes such processes as cognitive control, statistical learning, and working memory, we focus on the effects of aging on episodic memory processes including basic episodic memory, false memory, and associative memory.
False Memories
Recent Publications:
Chamberlain, J. D., Bowman, C. R., & Dennis, N. A. (2022). Age-related differences in encoding-retrieval similarity and their relationship to false memory. Neurobiology of Aging, 113, 15-27.
Chamberlain, J. D., Turney, I. C., Goodman, J. T., Hakun, J. G., & Dennis, N. A. (2021). Fornix white matter microstructure differentially predicts false recollection rates in older and younger adults. Neuropsychologia, 157, 107848.
Bowman, C. R., Chamberlain, J. D., & Dennis, N. A. (2019). Sensory representations supporting memory specificity: Age effects on behavioral and neural discriminability. Journal of Neuroscience, 39(12), 2265-2275.

While the vast majority of memory-related research has focused on factors underlying memory success, false memories, or memory for facts, items, or events that never happened, are equally as critical to understanding memory function. Critically, with respect to aging, research has shown that age-related memory errors arise as much from forgetting as they do from false memories. Examples of false memories include remembering that you took your morning medication when in fact you did not, or incorrectly identifying an innocent by-stander as the perpetrator of a crime. Thus, the impact of false memories can have potentially life-alternating consequences. Age-related increases in false memories have been attributed to a deficit in memory for item-specific information coupled with a reliance on memory for gist (semantic or perceptual features shared by a group of past events). Over the past decade our research has consistently linked this behavior to age deficits in visual cortex activity and activation in lateral temporal cortex that correlates with individual differences in memory performance (Dennis et al., 2007; 2008; 2014; Dennis & Turney, 2018; Webb & Dennis, 2018).
Our research also finds that, when successfully avoiding false memories, the cognitive and neural basis of correctly rejecting lures differs with respect to the relatedness between lures and studied items (Bowman & Dennis, 2015a,b; 2016). Specifically, correct rejections of lures that were related to studied items elicit activation in the hippocampus, visual and lateral prefrontal cortices, implicating the roles of memory retrieval, perceptual processing, and retrieval monitoring for the suppression of false memories. Conversely, we found that older adults did not differentially recruit the foregoing regions to successfully avoid memory errors, suggesting that they have difficulty using item-specific details and monitoring to distinguish between studied and unstudied information. Future work in this area aims to identify individual differences factors that contribute to false memories across the lifespan and to examine the effectiveness of strategy-based training aimed at reducing memory errors.
Associative Memories
Recent Publications:
Dennis, N. A., Overman, A. A., Carpenter, C. M., & Gerver, C. R. (2022). Understanding associative false memories in aging using multivariate analyses. Neuropsychology, development, and cognition. Section B, Aging, Neuropsychology, and Cognition, 29(3), 500-525. https://doi.org/10.1080/13825585.2022.2037500
Gerver, C. R., Overman, A. A., Babu, H. J., Hultman, C. E., & Dennis, N. A.(2020). Examining the neural basis of congruent and incongruent configural contexts during associate retrieval. Journal of Cognitive Neuroscience. 32(9): 1796-1812.
Overman, A.A., McCormick-Huhn, J.M., Dennis, N.A., Steinsiek, A.B., & Cesar, L.B. (2019). Same face, same place, different memory: Manner of presentation modulates the associative deficit in older adults. Aging, Neuropsychology, and Cognition. 26:1, 44-57.

Associative memory refers to memory for the relationship between two or more items or between an item and its context. As such, associative memory allows us to combine individual details of an event into a single complex memory. One common, real-world example of associative memory involves the everyday need to remember when and where you may meet someone. For example, on any given day you may experience many different events, such as being at work, at the grocery store, and at a cocktail party, during which time you may meet several new people in each context. Without the ability to form a link between the people and the location (i.e., to bind item and contextual information), it would be impossible to later recall who you met where. Aging research finds that age-related declines in associative memory exceed those of simply item memory, making associative deficits a critical issue affecting older adults.
Our previous work has shown that successful encoding activity in hippocampal and prefrontal regions is significantly attenuated in older adults during associative memory (e.g., study faces with a pictoral context – face/scene pairs) compared to that seen in item memory (e.g., studying faces and scenes separately). However, despite overall decreases in activation levels, functional connectivity between these two regions is enhanced in older adults. The functional connectivity findings are consistent with the posterior-anterior shift with aging (PASA) (Dennis et al., 2008). More recently we have shown that the manner of presentation influences associative memory performance in aging. Specifically, aging has a greater impact on item-context compared to item-item associations (Overman et al., 2018). Additional work in the lab suggests that item familiarization can enhance associative memory, also reducing the neural demands associated with binding processes (Dennis et al., 2015). Ongoing work continues to focus on the effects of aging as well as identifying means for mitigating age-related declines in associative memory.
Multivariate Approaches
https://github.com/canlabpsu?tab=repositories
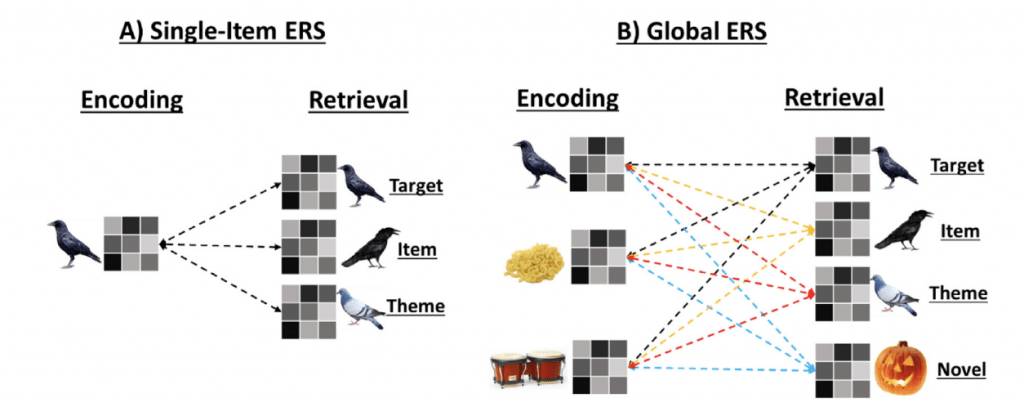
The lab also utilizes multivoxel pattern analyses (MVPA) and pattern similarity analyses, such as encoding-retrieval similarity (ERS) analysis, to investigate differences underlying neural patterns associated with memory performance in younger and older adults. These methods examine differences in the distributed pattern of brain activity rather than the amplitude of neural activity, allowing us to make interpretations as to the representations underlying episodic memories.
For example, with respect to our studies of false memories, we recently showed that portions of the ventral visual stream could discriminate neural patterns of target and lure items across multiple levels of similarity in both younger and older adults. Interestingly, the relationship between this neural discriminability and memory discriminability was age invariant in some visual regions, but was moderated by age in other regions. Results have implications for age-related memory deficits when old and new information share perceptual and semantic similarities (Bowman, Chamberlain, and Dennis, 2019). Additional work in progress is investigating whether cognitive training in older adults can increase neural discriminability within the retrieval network, leading to reduced false memory errors Chamberlain, Turney, Goodman, Hakun, & Dennis, 2021. Finally, using ERS, we have found that the similarity of neural patterns between encoding and retrieval is reduced in older compared to younger adults in visual and temporal regions, but that some of these differences can be accounted for by baseline neural similarity effects in older adults (Chamberlain, Bowman & Dennis, 2022).
We have also used multivariate analyses to investigate neural activation patterns underlying different types of associative memory in both young and older adults. In one study we found that during associative encoding, neural patterns in both occipital and frontal regions were discriminable with respect to whether the stimuli were presented as an item-item or an item-context association (Dennis, Overman, et al., 2022). While in a second study we showed that neural patterns associated with associative memory were also sensitive to whether individual items comprising associations were previously encountered or not (Elbich, Webb, & Dennis, 2021).